
実用酵母への外来遺伝子の導入および不要ベクター配列の除去
pAUR135 DNA (製品コード 3604)
二倍体である焼酎酵母の一方のADE2遺伝子内に、pAURを用いてアルコール脱水素酵素ADH1のプロモーターに連結したlacZ遺伝子を挿入して、不要配列を除去して発現させることを試みた。
図1. 実験の概要およびプライマー(P1~P4)の位置
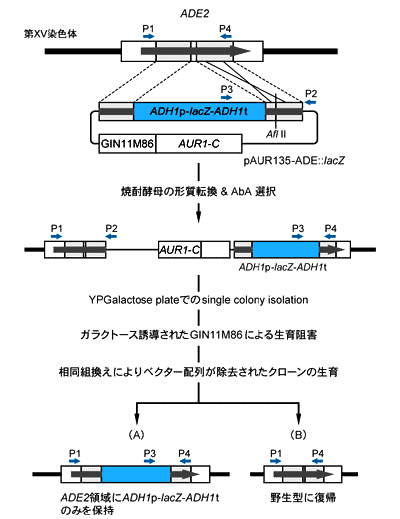
- (1)形質転換用プラスミドの構築
- 焼酎酵母協会2号のゲノムDNAを鋳型としたPCRにより、ADE2遺伝子の一部領域、約1.6 kbpを増幅し、このDNA断片をpAUR135のSma I-EcoR Iサイトに挿入してプラスミドpAUR135-ADEを作製した。次に、pAUR123-lacZ(pAUR123にlacZ遺伝子を組み込んだもの)を鋳型としたPCRを行い、ADH1p-lacZ-ADH1t領域、約4.1 kbpを増幅した。これをpAUR135-ADEのADE2断片上のHind III-Bgl IIサイトに連結して、形質転換用のプラスミドpAUR135-ADE::lacZを構築した。
- (2)焼酎酵母の形質転換
- pAUR135-ADE::lacZをADE2断片上のAfl IIサイトで1ヶ所切断して直鎖状にし、そのプラスミド5 μgを用いて酢酸リチウム法で焼酎酵母の形質転換を行った。YPD液体培地で一晩培養した後、0.5 μg/mlのAureobasidin A(AbA)を含むYPD選択プレートで形質転換体を選択した。約102/μg DNAの形質転換効率で、形質転換体が得られた。これらの形質転換体はいずれもAbA耐性とβ-ガラクトシダーゼ活性(0.15~0.25 U/OD600)を有していた。
- (3)ベクター除去クローンの選別
- (2)で得た形質転換体、4クローンをそれぞれYPGalactoseプレート上にストリークした。ガラクトース誘導によってGIN11M86が高発現されて明らかな生育阻害が起こったことを確認するとともに、十数個ずつのシングルコロニーを分離した。これらの単離クローンはいずれもAbA感受性であり、相同組換えにより不要なpAUR135ベクター配列が除去されて生じたコロニーと考えられる。
- (4)lacZ保持クローンのスクリーニング
- (3)で得たベクター配列除去クローンから、図1(A)の相同組換えが起こり、lacZ遺伝子がADE2遺伝子中に保持されているクローンをスクリーニングするために、o-Nitrophenyl-β-D-Galactopyranoside(OPNG)を基質として各クローンのβ-ガラクトシダーゼ活性を調べた。その結果、各形質転換体から得たベクター除去クローンのうちの0~20%で、形質転換体と同等ないしやや低めのβ-ガラクトシダーゼ活性(0.05~0.2 U/OD600)が認められた。
- (5)lacZ遺伝子のゲノムへの組込みおよびベクター配列除去の確認
- 焼酎酵母協会2号、(2)で得た形質転換体2株、(3)~(4)で得たlacZ遺伝子を保持しているベクター除去クローン2株、および図1(B)の相同組換えが起こり野生型に戻ったと考えられるクローン2株から、それぞれGenとるくん(酵母・グラム陽性菌用)(製品コード 9083)を用いてゲノムDNAを調製した。これらのゲノムDNAを鋳型とし、図1に示したプライマー(P1~P4)を組み合わせてPCRを行い、ゲノム上のADE2遺伝子領域へのpAUR135-ADE::lacZの組込み[図2(1)のレーン3, 4]、pAUR135ベクター配列の除去[図2(1)のレーン5~8]、ベクター除去後のlacZ遺伝子の保持[図2(2)のレーン5, 6]を確認した。
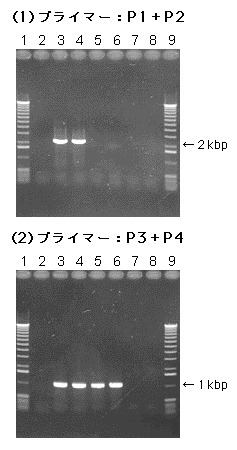 レーン
レーン
1, 9:DNAサイズマーカー
2:焼酎酵母協会2号
3, 4:pAUR135-ADE::lacZによる形質転換体
5, 6:lacZを保持したベクター除去クローン
7, 8:野生型に戻ったベクター除去クローン
図2. PCRによる、ゲノムへの組込みおよびベクター除去の確認